Книга: Новая наука о жизни
Навигация: Начало Оглавление Другие книги
7.4. Доминирование
На первый взгляд может показаться, что идея о стабилизации видовых форм путем морфического резонанса от прошлых организмов того же вида мало что добавляет к обычному объяснению в рамках одного лишь генетического подобия. Однако важность этой идеи становится очевидной при рассмотрении гибридных организмов, которые подвержены морфическому резонансу от двух различных родительских типов.
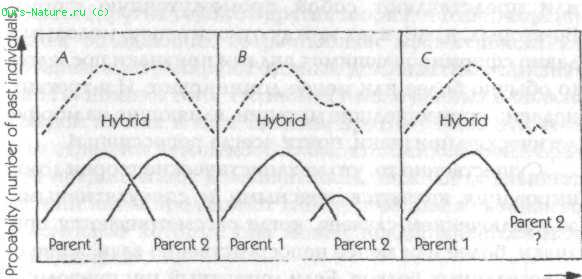
Рис. 19. Диаграмма вероятностных структур морфогенетических полей родителей и гибридов
Вернемся к аналогии с радио: при нормальных обстоятельствах приемник настроен в любой момент времени только на одну станцию, так же как организм в норме «настроен» на прошлые подобные системы того же вида. Но если радио должно быть настроено на две станции одновременно, то звуки, которые оно издает, зависят от относительной силы сигналов этих станций: если один очень сильный, а второй очень слабый, то эффект последнего мало заметен; но если оба одинаково сильны, приемник издает смесь звуков из обоих источников. Подобно этому, в гибриде, образовавшемся в результате скрещивания двух видов, присутствие генов и характерных для каждого вида генных продуктов будет стремиться привести развивающийся организм в состояние морфического резонанса с прошлыми организмами обоих родительских типов. Тогда полный набор вероятностных структур в пределах морфогенетических полей гибрида будет зависеть от относительной силы морфического резонанса двух родительских типов. Если оба родителя происходят из видов, представленных сравнимым числом прошлых индивидуумов, то оба будут влиять на морфогенез в одинаковой степени, давая комбинацию или «результирующую» двух родительских форм (рис. 19 А). Но если один вид был представлен меньшим числом индивидов, чем другой, меньший вклад первого в общую вероятностную структуру будет означать, что станет преобладать форма второго из родительских видов (рис. 19 В). А если один из родителей происходит из недавно образовавшейся мутантной линии, морфический резонанс от малого числа прошлых индивидуумов этого типа внесет лишь незначительный вклад в вероятностную структуру гибридов (рис. 19 C).
Эти ожидания согласуются с фактами. Во‑первых, гибриды между двумя хорошо утвердившимися разновидностями или видами обычно сочетают черты обоих или представляют собой промежуточную форму. Во‑вторых, в гибридах между относительно молодым и давно сформировавшимся видами признаки последнего обычно более или менее доминируют. И в‑третьих, недавно произошедшие мутации, влияющие на морфологические признаки, почти всегда рецессивны.[150]
Существенно то, что механистические теории доминирования являются неясными и спекулятивными, за исключением случаев, когда рассматриваются признаки, более или менее непосредственно зависящие от определенных белков. Если мутантный ген приводит к потере функции, например стимулируя синтез дефектного фермента, то он будет рецессивным, поскольку в гибридах присутствие нормального гена делает возможным производство нормального фермента и, следовательно, протекание нормальных биохимических реакций. Однако в некоторых случаях продукт дефектного гена может быть опасным, например нарушая проницаемость мембран, и тогда мутация будет одновременно доминирующей и летальной.
Эти объяснения удовлетворительны в пределах своей применимости; но в отсутствие какого‑либо механистического понимания морфогенеза попытка объяснить доминирование в наследовании формы путем экстраполяции с молекулярного уровня неизбежно приводит к необходимости считать этот спорный вопрос решенным и не требующим доказательств.
Принятые генетические теории доминирования более сложные, нежели чисто биохимическая теория; они подчеркивают, что доминирование не фиксировано, но оно эволюционирует. Чтобы объяснить относительную однородность диких популяций, в которых большинство нелетальных мутаций рецессивны, они допускают, что доминированию «дикого типа» благоприятствует естественный отбор. Одна теория постулирует отбор генов, которые модифицируют доминирование других генов,[151] а другая теория – отбор версий генов, обладающих возрастающей эффективностью, которые контролируют данные доминантные признаки.[152] Но помимо того, что имеется мало данных в пользу каждой из них и есть таковые против обеих, эти теории страдают тем недостатком, что они, скорее, заранее предполагают доминантность, нежели объясняют ее: они лишь предлагают гипотетические механизмы, с помощью которых она может поддерживаться или усиливаться.[153]
Согласно гипотезе формативной причинности, доминантность развивается по причине фундаментально иного характера. Типы, которым благоприятствует естественный отбор, будут представлены большим числом индивидуумов, чем типы с меньшей способностью к выживанию; с течением времени первые приобретают возрастающую доминантность в результате кумулятивного эффекта морфического резонанса.
Эту гипотезу в принципе можно экспериментально отличить от всех механистических теорий доминантности. Согласно последним, при данных условиях окружающей среды доминантность зависит только от генетического строения гибрида, тогда как, согласно первой, она зависит как от генетического строения, так и от морфического резонанса родительских типов. Поэтому, если соотношение сил резонанса от родительских типов изменяется, то доминирование одного над другим также изменится, даже если генетическое строение гибрида останется тем же.
Рассмотрим следующий эксперимент. Гибридные семена получены скрещиванием между растениями четко определенного сорта (Р1) и мутантной линии (Р2). Некоторые из этих гибридных семян помещены в холодный инкубатор, а другие выращиваются в контролируемых условиях. Характеристики гибридных растений тщательно регистрируются, и сами растения сохраняются. В этих растениях абсолютно доминирует морфология Р1 (рис. 19 С). Затем очень большие количества растений мутантного типа (Р2) выращиваются в поле. Некоторые из полученных затем гибридов снова выращиваются в тех же условиях, что и раньше, из той же партии семян. Поскольку теперь Р2 вносит больший вклад путем морфического резонанса, Р1 может доминировать лишь отчасти (рис. 19 В). После нового выращивания множества растений Р2 форма гибрида может стать промежуточной между двумя родительскими типами (рис. 19 А). Затем выращивается еще больше растений Р2; после этого гибриды снова выращиваются в тех же условиях, что и предыдущие из той же партии семян. Теперь тип Р2 будет вносить путем резонанса больший вклад, чем Р1 и морфология Р2 станет доминантной.
Такой результат явится весомым свидетельством в пользу гипотезы о роли морфического резонанса в доминировании признаков и будет абсолютно непонятным с точки зрения ортодоксальной генетической теории. Единственная проблема в таком эксперименте состоит в том, что его может быть трудно осуществить на практике, поскольку если Р1 – это хорошо определенный тип, который уже существовал очень долгое время – в случае дикого типа, возможно, тысячи или даже миллионы лет, – то практически невозможно вырастить сравнимые количества типа Р2. Эксперимент может быть осуществим, только если P1 – это недавно появившийся сорт, у которого количество особей, выращенных в прошлом, было сравнительно невелико.
7.3. Измененные пути морфогенеза7.5. Семейные сходства