Книга: Новая наука о жизни
Навигация: Начало Оглавление Другие книги
11.2. Морфический резонанс и поведение: экспериментальная проверка
В механистической биологии проводится резкое различие между врожденным и обученным поведением: предполагается, что первое «генетически запрограммировано», или «закодировано», в ДНК, тогда как последнее рассматривается как результат физико‑химических изменений в нервной системе. Невозможно представить, каким способом эти изменения могут специфически модифицировать ДНК (такое требование выдвигали бы последователи ламаркизма); поэтому считается невозможным, чтобы обученное поведение, приобретенное животным, могло наследоваться его потомством (конечно, исключая «культурное наследование», при котором потомство воспринимает модели поведения от своих родителей).
В противоположность этому, согласно гипотезе формативной причинности, нет качественного различия между врожденным и обученным поведением, поскольку оба зависят от моторных полей, даваемых морфическим резонансом (раздел 10.1). Поэтому эта гипотеза допускает возможную передачу обученного поведения от одного животного к другому и приводит к проверяемым предсказаниям, которые отличаются не только от таковых ортодоксальной теории наследования, но также и от предсказаний ламаркизма.
Рассмотрим следующий эксперимент. Дикие животные помещаются в условия, где они учатся реагировать на данный стимул неким характерным образом. Затем их заставляют повторять эту модель поведения много раз. Согласно гипотезе, новое моторное поле будет усилено морфическим резонансом, который не только приведет к тому, что поведение обученных животных станет все более привычным, но и повлияет так же, хотя и менее специфично, на других животных, подвергаемых тому же стимулу: чем больше будет число животных, научившихся выполнять данное задание в прошлом, тем легче его будет выучить последующим подобным же животным. Поэтому в экспериментах такого типа должна быть возможность наблюдать прогрессивное увеличение скорости обучения не только у животных, происходящих от обученных предков, но также у генетически подобных животных, происходящих от необученных предков. Это предсказание отличается от такового ламаркистской теории, согласно которой только потомки обученных животных должны обучаться быстрее. А по общепринятой теории не должно наблюдаться увеличение скорости обучения потомков как необученных, так и обученных животных.
Сказанное можно суммировать следующим образом: повышенная скорость обучения в последовательных поколениях как тренированных, так и нетренированных линий подтверждала бы гипотезу формативной причинности: увеличение скорости только в тренированных линиях – ламаркизм, а отсутствие такого увеличения в обоих линиях – ортодоксальную теорию.
Эксперименты этого типа фактически уже проводились. Результаты подтверждают гипотезу формативной причинности.
Оригинальный эксперимент был начат в Гарварде в 1920 году У. Мак‑Дугаллом, который надеялся провести тщательную проверку возможности наследования приобретенных особенностей поведения, предполагаемых ламаркизмом. Экспериментальными животными были белые крысы уистарской (Wistar) линии, которые – бережно выращивались в лабораторных условиях в течение многих поколений. Их задача состояла в том, чтобы научиться выбираться из специально сконструированного бака с водой, доплывая до одного из двух проходов, из которого можно было выйти наружу. Ложный проход был ярко освещен, тогда как истинный проход не освещался. Если крыса выходила через освещенный проход, она получала удар электрошоком. Два прохода освещались по очереди, один раз – первый, следующий раз – второй. За меру скорости обучения принималось число ошибок, которые допускала крыса, прежде чем она усваивала, что выходить надо через неосвещенный проход:
«Некоторым крысам требовалось целых 330 погружений, причем приблизительно половина из них сопровождалась электрошоком, прежде чем они научались избегать освещенный проход. Во всех случаях процесс обучения внезапно достигал критической точки. В течение длительного времени животное выказывало явное отвращение к освещенному проходу, часто колебалось, прежде чем войти в него, поворачивало назад или отчаянно бросалось внутрь; но, не уловив постоянной связи между ярким светом и шоком, оно продолжало устремляться в этот проход столь же часто, как и в другой. Наконец в обучении наступал момент, когда животные, оказавшись перед освещенным проходом, определенно и решительно поворачивали обратно, искали другой, смутно видимый проход и спокойно вылезали наружу. После достижения этой точки в обучении ни одно животное не делало ошибку, снова выбирая яркий проход, разве что в очень редких случаях» .[229]
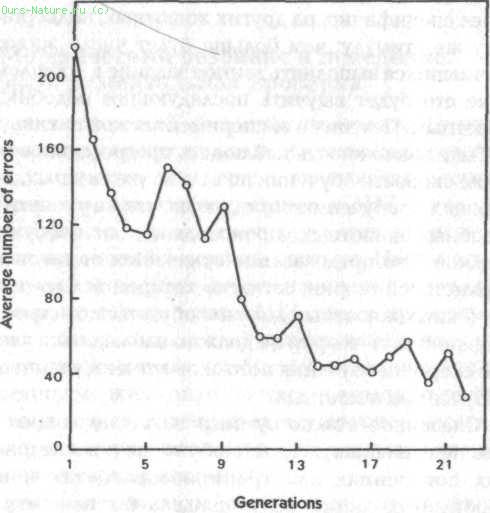
Рис. 28. Среднее число ошибок у последовательных поколений крыс, отобранных в каждом поколении по медленности обучения (данные из McDougall, 1938)
В каждом поколении крысы, от которых должно было быть выведено следующее поколение, отбирались случайным образом до того, как определялась скорость их обучения, тогда как спаривание происходило после их тестирования. Такая процедура была выбрана для того, чтобы избежать любой возможности сознательного или неосознанного отбора в пользу более быстро обучающихся крыс.
Этот эксперимент продолжался на протяжении 32 поколений крыс в течение 15 лет. В согласии с теорией Ламарка в последовательных поколениях крыс наблюдалась заметная тенденция к увеличению скорости обучения. Об этом свидетельствовало среднее число ошибок, которые делали крысы: в первых восьми поколениях оно превышало 56, а во второй, третьей и четвертой группах из восьми поколений – соответственно, 41, 29 и 20.[230] Разница была очевидной не только в количественных данных, но также в фактическом поведении крыс, которые в последующих поколениях становились более осторожными и опытными.[231]
Мак‑Дугалл предвидел критику того рода, что, несмотря на случайный отбор родителей в каждом поколении, все же мог вкрасться какой‑то отбор в пользу более быстро обучающихся крыс. Чтобы проверить такую возможность, он начал новый эксперимент с другой группой крыс, в которой родители действительно отбирались на основе очков, полученных ими в процессе обучения. В одной серии выбирались только быстро, в другой – только медленно обучающиеся крысы. Как и ожидалось, потомство быстро обучающихся крыс обучалось сравнительно быстро, а потомство медленно обучающихся – сравнительно медленно. Однако даже в последней серии обучение у последних поколений заметно улучшалось, несмотря на повторяющийся отбор в пользу медленно обучающихся крыс {рис. 28).
Эти эксперименты проводились весьма тщательно, и критики не могли отвергнуть результаты по причине технических изъянов. Но они привлекли внимание к недостаткам в постановке экспериментов: Мак‑Дугаллу не удалось осуществить систематическую проверку изменения скорости обучения крыс, родители которых не проходили обучения.
Один из этих критиков, Ф. А. Е. Крю из Эдинбурга, повторил эксперименты Мак‑Дугалла с крысами, происходившими от той же инбридинговой линии,[232] используя бак подобной же конструкции. Он включил также параллельную линию «нетренированных» крыс, отдельные из которых тестировались в каждом поколении на скорость обучения, тогда как другие, которые не тестировались, рождали потомство. Через 18 поколений в таком эксперименте Крю не обнаружил систематического изменения в скорости обучения ни в тренированной, ни в нетренированной линиях.[233] Вначале казалось, что это порождает серьезные сомнения в результатах Мак‑Дугалла. Однако эти две группы результатов нельзя было непосредственно сравнивать в силу трех обстоятельств. Во‑первых, по какой‑то причине крысы гораздо легче обучались в опытах Крю, чем в более ранних поколениях Мак‑Дугалла. Этот эффект был столь сильно выражен, что значительное число крыс как в тренированных, так и в нетренированных линиях «решали» задачу немедленно, не получая ни единого шока! Средние значения очков у крыс в опытах Крю с самого начала были примерно такими же, как у крыс Мак‑Дугалла через более чем 30 поколений тренировки. Ни Крю, ни Мак‑Дугалл не могли дать удовлетворительного объяснению этому несоответствию. Но, как указывал Мак‑Дугалл, поскольку цель исследования состояла в том, чтобы выявить любые воздействия тренировки на последующие поколения, эксперимент, в котором некоторые крысы не тренировались вообще, а многие другие – очень мало, нельзя было рассматривать как способный продемонстрировать такое воздействие.[234] Во‑вторых, в результатах Крю обнаруживались большие и кажущиеся случайными флуктуации от поколения к поколению, гораздо большие, чем флуктуации в результатах Мак‑Дугалла, и эти большие флуктуации вполне могли скрыть любую тенденцию к улучшению очков в более поздних поколениях. В‑третьих, Крю выбрал путь очень жесткого инбридинга (скрещивания очень близких родственников), скрещивая в каждом поколении братьев с сестрами. Он не ожидал, что это даст неблагоприятный эффект, поскольку крысы происходили от одной главней инбридинговой семьи, с которой все начиналось:
«Даже история моей главной семьи выглядит как эксперимент по инбридингу. Имеются широкая основа семейных линий и узкий кончик двух оставшихся линий. Скорость воспроизведения падает, и угасает одна линия за другой» .[235]
Даже в выживающих линиях многие животные рождались со столь серьезными аномалиями, что их приходилось отбрасывать. Вредные последствия такого жесткого инбридинга вполне могли маскировать любую тенденцию к увеличению скорости обучения. В целом эти дефекты эксперимента Крю означают, что его результаты можно рассматривать только как предварительные, и сам он фактически признавал, что вопрос остается открытым.[236]
К счастью, эта история здесь не кончается. Эксперимент был снова проведен У. Е. Эгером и его коллегами в Мельбурне с использованием методов без тех недостатков, которые были у Крю. На протяжении 20 лет они измеряли скорости обучения тренированных и нетренированных линий в 50 последовательных поколениях. В согласии с данными Мак‑Дугалла они обнаружили, что у крыс тренированной линии существует явная тенденция к более быстрому обучению в последующих поколениях. Но точно такая же тенденция была обнаружена и в нетренированной линии .[237]
Можно удивляться, почему же Мак‑Дугалл не наблюдал подобный эффект на своих нетренированных линиях? Ответ состоит в том, что он его наблюдал.
Несмотря на то что он тестировал контрольных крыс, происходивших от исходной главной семьи лишь изредка, он заметил «неприятный факт, что группы контрольных крыс, произведенных от этой семьи в 1926, 1927, 1930 и 1932 годах, демонстрировали уменьшение среднего числа ошибок с 1927‑го по 1932 год». Он думал, что этот результат мог быть случайным, но добавил следующее:
«Возможно, что выпадение по среднему числу ошибок периода с 1927‑го по 1932 год выражает реальное изменение конституции всей семьи, ее улучшение (относительно этой определенной способности), природу которого я не могу предположить» .[238]
С публикацией окончательного отчета группы Эгера в 1954 году закончилась затянувшаяся полемика по поводу «ламаркистского эксперимента Мак‑Дугалла». Одинаковое улучшение результатов как в тренированной, так и в нетренированной линиях исключало интерпретацию в духе ламаркизма. Вывод Мак‑Дугалла был отвергнут, и это, казалось, исчерпывало вопрос. С другой стороны, его результаты получили подтверждение.
Эти результаты казались совершенно необъяснимыми; они были бессмысленными с точки зрения какой‑либо из действующих концепций и никогда не были доведены до конца. Но они вполне осмысленны в свете гипотезы формативной причинности. Конечно, сами по себе они не могут доказать справедливость этой гипотезы; всегда можно предложить другие объяснения, например то, что последовательные поколения крыс становились все более разумными по неизвестной причине, не связанной с их тренировкой.[239]
В будущих экспериментах наиболее прямым способом проверки действия морфического резонанса, вероятно, будет такой, в котором большое число крыс (или любых других животных) обучается новой задаче в одном месте, а затем такие же крысы учатся выполнять ту же задачу в другом месте, за сотни миль от первого, и будет выясняться, есть ли увеличение в скорости обучения у этих последних. Начальная скорость обучения в обеих группах должна быть более или менее одинакова. Тогда, согласно гипотезе формативной причинности, скорость обучения должна прогрессивно возрастать в том месте, где тренируется большое число животных; и такое же увеличение скорости должно обнаруживаться также в другом месте, даже несмотря на то что здесь число испытуемых крыс очень мало. Очевидно, следует принять меры предосторожности, чтобы избежать любых возможных сознательных или неосознанных предубеждений со стороны экспериментаторов. Один из способов может состоять в том, чтобы во втором месте тестировать скорости обучения крыс, давая им несколько разных задач с правильным интервалом, например раз в месяц. Затем в первом месте выбирается наугад из этого набора определенная задача, которой обучаются тысячи крыс. Более того, время начала обучения также будет выбрано наугад, например четыре месяца спустя после начала выполнения набора задач во втором месте. Экспериментаторы во втором месте не будут знать ни того, какая задача была выбрана, ни того, когда началось обучение ей в первом месте. Если при этих условиях во втором месте будет обнаружено заметное увеличение в скорости обучения данной задаче после того, как обучение началось в первом месте, этот результат будет весомым подтверждением справедливости гипотезы формативной причинности.
Эффект такого типа вполне мог иметь место, когда группы Крю и Эгера повторяли эксперимент Мак‑Дугалла. В обоих случаях их крысы с самого начала обучались своей задаче значительно быстрее, чем это делали крысы Мак‑Дугалла, когда он впервые начал свой эксперимент.[240]
Если предложенный выше эксперимент действительно был бы осуществлен и дал бы положительные результаты, он не мог бы быть полностью воспроизводим по самой своей природе, ибо при попытках его повторить на крыс должен будет действовать морфический резонанс от крыс в первоначальном эксперименте. Чтобы продемонстрировать тот же эффект снова и снова, было бы необходимо изменить либо задачу, либо вид, тестируемый в каждом эксперименте.
11.1. Наследование поведения11.3. Эволюция поведения