Книга: Общая экология
Навигация: Начало Оглавление Другие книги
8.6.4. Механизмы динамики численности
Типы популяционной динамики численности наиболее хорошо изучены у лесных насекомых, на примере которых выявляется большое многообразие конкретных связей и особенностей видов, влияющих на плотность их популяций.
В лесных сообществах преобладают виды со стабильным ходом популяционной динамики. Они питаются практически за счет всех частей деревьев (стволы, корни, побеги, листья, почки, плоды, семена), однако при этом не оказывают губительного воздействия на растения. Как правило, такие индифферентные виды отличаются низкой миграционной активностью, ведут оседлый образ жизни. Комплекс их паразитов включает обычно несколько сильно специализированных видов, эффективно поражающих популяцию хозяина.
Флюктуирующий тип динамики численности свойствен многим ксилофагам (потребителям коры и древесины): усачам, златкам, короедам. Для них характерно совместное заселение кормовых объектов – ослабление деревьев. Это позволяет быстро снизить устойчивость дерева, но в то же время совместное обитание ксилофагов обостряет конкурентные отношения между ними, что действует как безынерционный механизм регуляции численности.
Для короедов, поселяющихся первыми на ослабленных деревьях, при избыточном запасе питательных веществ (растворимых углеводов и крахмала) развитие лимитируется воздействием защитных реакций дерева, таких, например, как смоловыделение у хвойных. Кроме того, еще отсутствуют симбиотические микроорганизмы. Сопротивление еще жизнеспособных деревьев может быть сломлено при концентрированном нападении и избыточно высокой плотности вредителя. Другой предел пригодности дерева для короедов – полное отмирание и разрушение луба. Между двумя указанными состояниями дерева для короедов создается оптимум кормовых условий, отмечается максимальная выживаемость всех фаз развития и наиболее высокие коэффициенты размножения.
Относительную роль различных регуляторных механизмов, определяющих флюктуирующий ход численности лесных насекомых, можно проследить на примере большого лиственного короеда – Ips subelongatus. Этот вредитель лиственницы и частично других пород широко распространен по всей таежной зоне.
Жуки нападают на физиологически ослабленные живые лиственницы, у которых отсутствует активное защитное смоловыделение. Внедряясь под кору, самцы продуцируют запаховые вещества – феромоны, привлекающие других особей, чем обеспечивается массовость заселения подходящего дерева и его дальнейшее ослабление. Феромоны играют также роль химических регуляторов внутрипопуляционных отношений – распределения короедов по стволу, своего рода меткой территории отдельных гнезд. Жуки, не нашедшие места на данном дереве, переселяются на другие. В дальнейшем внутривидовая конкуренция регулируется самками, которые в зависимости от плотности поселения изменяют длину маточных ходов и количество откладываемых яиц. При возрастающем недостатке площади часть самцов и самок мигрирует на другие объекты. Личинка короеда за период развития потребляет 0,78 см6 луба. Если «кормовые площадки» личинок при высокой плотности поселения становятся меньше этого предела, среди них увеличивается смертность. Таким образом, внутривидовая конкуренция регулирует заселенность дерева на всех фазах жизни вредителя: на стадии взрослых жуков она ведет к усилению миграций, на стадии личинок – к гибели части популяции. Показатели размножения остаются высокими на всех этапах флюктуации численности популяции.
Естественных врагов данного вредителя можно разделить на пять групп, по особенностям их взаимодействия с короедом. 1. Враги, действующие в период лёта короедов, – птицы (синицы, дятлы и др.), ктыри, муравьи. Они уничтожают часть популяции, но одновременно и энтомофагов короеда, т. е. других его естественных врагов. Их регуляторная деятельность поэтому слабо эффективна. 2. Хищные насекомые, уничтожающие короеда на всех фазах развития под корой (ряд пестрянок, чернотелок, карапузиков, хищных мух и клопов). При низкой и средней плотности вредителя хищники сдерживают рост его численности, при высокой же плотности короедов – способствуют повышению выживаемости их вследствие ослабления внутривидовой конкуренции. 3. Паразиты, заражающие только личинок короедов, – бракониды, хальциды. На регуляцию численности воздействуют слабо, так как пораженные личинки погибают лишь в конце жизни и общая смертность их от паразитов составляет несколько процентов. 4. Паразиты, заражающие взрослых жуков, – нематоды, некоторые перепончатокрылые. Их влияние проявляется в снижении плодовитости и сокращении продолжительности жизни жуков, регулирующий эффект в целом невысок. 5. Бактериальные и грибные заболевания. Эпизоотии среди короедов возникают лишь при резком ухудшении условий, когда действуют более мощные механизмы снижения численности.
Таким образом, в межвидовых отношениях короедов более или менее значительную роль в регуляции их численности имеют лишь хищные насекомые при относительно невысокой численности вредителя.
Способность к вспышкам массового размножения свойственна относительно немногим видам лесных насекомых. В сибирской тайге у скрытоживущих видов взрывной тип динамики численности характерен для алтайского лиственничного усача, большого черного хвойного усача, лиственничной почковой галлицы и некоторых других. Среди открытоживущих листогрызущих насекомых способность давать вспышки массового размножения свойственна лишь некоторым видам чешуекрылых и перепончатокрылых (пилильщикам, ткачам). Отличительные черты экологии таких видов: высокая выживаемость в сильно изменчивой среде благодаря специальным адаптациям, высокой миграционной активности, высокой и изменчивой плодовитости. У открытоживущих видов часто выявляется эффект группы и фазовая изменчивость.
Один из опаснейших вредителей хвойных пород – сибирский шелкопряд Dendrolimus sibiricus, распространенный от Урала до Тихого океана. Этот вид обладает следующими внутрипопуляционными механизмами регуляции численности.
В очагах сибирского шелкопряда по фазам вспышки существенно изменяется соотношение полов. Доля самок варьирует от 32 до 76 %. При нарастании вспышки доминируют самки, при затухании – самцы. В переуплотненных популяциях повышается смертность самок на всех фазах развития, а также отмечается их более высокая миграционная активность из очагов размножения. В фазе максимума численности на периферии очага доля самок составляет до 73 %, а в центре – 44 %.
У сибирского шелкопряда, как и у других видов с вспышками массового размножения, плодовитость изменяется в широких пределах. Этот показатель сильно зависит от плотности популяции (рис. 140). При низкой и сверхвысокой численности плодовитость самок снижается почти втрое. Наиболее высокая плодовитость наблюдается в фазе нарастания численности (свыше 300 яиц на самку), а затем закономерно снижается под влиянием конкуренции и недостатка корма в переуплотненных очагах. Изменение соотношения полов и плодовитости можно рассматривать как безынерционные или слабоинерционные механизмы регуляции численности. Их действие проявляется в течение жизни одного поколения.
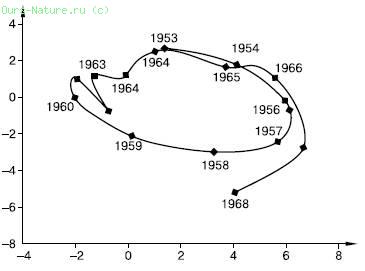
Рис. 140. Фазовый портрет динамики численности сибирского шелкопряда в Красноярском крае (из А. С. Исаева и др., 2001):
по вертикали – логарифм коэффициента размножения;
по горизонтали – логарифм плотности популяции гусениц на одно дерево. Схема иллюстрирует цикличность вспышек и зависимость показателей размножения от плотности популяции
Миграционная активность сибирского шелкопряда – также эффективный внутрипопуляционный механизм регуляции численности. У этого вида отмечены разные типы массовых перемещений. В период низкой численности пульсирующие миграции перераспределяют вредителя на площади лесных массивов, в результате чего формируются резервации – места сохранения вида. В основном это перестойные пихтовые леса на плакорах или склонах южной экспозиции. В начальной фазе вспышки происходит стягивание вредителя с больших территорий на ограниченный участок леса с наиболее благоприятными условиями (конденсирующие миграции). К концу фазы нарастания численности начинается вылет бабочек из первичных очагов, где к этому времени плотность популяции достигает критических величин. На фазе максимума численности происходят массовые выселения бабочек и их перемещение воздушными потоками на расстояния до 100 км. Среди мигрантов преобладают самки с запасом зрелых яиц. В редких случаях отмечаются активные массовые миграции бабочек. При затухании очагов сибирского шелкопряда развиваются дисперсионные миграции – рассеивание популяции из очагов размножения в окружающие лесные массивы.
Для сибирского шелкопряда известен также эффект группы – положительное влияние оптимальной плотности, которое проявляется в основном у гусениц I–III возрастов и выражается в ускорении их развития более чем в 2 раза. В регуляцию хода численности вредителя вносит вклад также гетероцикличность – наличие особей с одно– и двухлетним циклом в зависимости от плотности популяции.
Повышение плотности очага сильно ускоряет метаболические процессы у членов популяции, что выражается как в ускорении сроков развития, так и в изменении поведения и внешнего вида бабочек и гусениц – появляются особи с темной окраской. В разных фазах цикла массового размножения количество темных бабочек в лесах Средней Сибири изменяется от 10 до 80 %.
Среди естественных врагов сибирского шелкопряда отмечено более 60 видов. Однако, несмотря на разнообразие, энтомофаги не могут постоянно держать под контролем численность вредителя из‑за высокой инерционности их количественной реакции. Усложняющий фактор – высокая активность сверхпаразитов. Многие энтомофаги сибирского шелкопряда гораздо менее его устойчивы к факторам абиотической среды. Например, один из эффективных паразитов яйцеед‑теленомус в массе гибнет в холодные зимы. Вот почему вспышки размножения сибирского шелкопряда часто возникают после холодных малоснежных зим.
При низкой численности сибирского шелкопряда в резервациях за счет естественных врагов‑энтомофагов гибнет до 95 % яиц, 40 % гусениц и 70 % куколок. Основными регуляторами при этом являются яйцеед‑теленомус и тахина‑мазицера. Общие показатели смертности в популяциях достигают более 99 %. Это свидетельствует о высоких возможностях вредителя переходить к вспышке. Быстрый рост численности может быть обеспечен даже небольшим (5‑10 %) увеличением выживаемости. На фазе нарастания численности поражаемость от энтомофагов падает (яйца – до 50 %, гусеницы – до 11 %, куколки – до 23 %). Энтомофаги «не успевают» сдерживать растущую популяцию вредителя. В период кульминации вспышки пресс энтомофагов на популяцию шелкопряда усиливается и достигает наибольшего выражения в период разреживания и депрессии численности, когда естественные враги полностью уничтожают избыток особей.
Таким образом, при массовых вспышках размножения лесные насекомые‑вредители выходят из‑под межвидового контроля и их возврат к стабильной численности происходит в основном в результате внутрипопуляционных регулирующих механизмов, которые наиболее эффективно действуют лишь при очень высоких уровнях плотности, губительной для лесного насаждения.
Понимание и учет всех этих факторов, определяющих динамику численности лесных вредителей, совершенно необходимы для принятия действенной системы мер борьбы с ними и профилактики вспышек, для определения необходимости, сроков, доз и форм применения средств, сдерживающих массовое размножение. С экологической точки зрения задача заключается в удержании популяции в зоне депрессии численности.
Понимание механизмов, определяющих движение численности популяций, дает в руки ключ управления ими. Однако эта задача не может решаться трафаретно для разных видов. Во всех случаях необходимо детальное знание экологических особенностей конкретного вида и условий его биотического окружения, а также характер взаимодействия модифицирующих факторов среды.
8.6.3. Типы динамики численности популяций8.6.5. Влияние ослабления или усиления пресса хищников на динамику популяций и структуру сообществ