Книга: Общая экология
Навигация: Начало Оглавление Другие книги
10.3. Стабильность биосферы
Основой самоподдержания жизни на Земле являются биогеохимические круговороты. Процессы созидания органического вещества, аккумулирующего энергию, и противоположные процессы его разложения с высвобождением этой энергии одинаково необходимы для существования жизни. Все доступные для живых организмов химические соединения в биосфере имеют предел. Исчерпаемость пригодных для усвоения химических веществ часто тормозит развитие тех или иных групп организмов в локальных участках суши или океана. По выражению академика В. Р. Вильямса, единственный способ придать конечному свойства бесконечного – это заставить его вращаться по замкнутой кривой. Все химические элементы, используемые в процессах жизнедеятельности организмов, совершают постоянные перемещения, переходя из живых тел в соединения неживой природы и обратно. Возможность многократного использования одних и тех же атомов делает жизнь на Земле практически вечной при условии постоянного притока нужного количества энергии.
Глобальный биогеохимический круговорот вещества на планете представляет собой систему сложно переплетенных циклов отдельных элементов, связанных друг с другом по аналогии с деталями часового механизма (рис. 163). Объединяясь в разных пропорциях в составе сложных органических молекул, синтезируемых в живых телах, атомы различных элементов вне живого вещества имеют свою «судьбу» в атмосфере, гидросфере и литосфере. Круговороты планетарного масштаба создаются из бесчисленных локальных циклических перемещений атомов, движимых жизнедеятельностью организмов в отдельных экосистемах, и тех перемещений, которые вызываются действием ландшафтных и геологических причин (поверхностный и подземный сток, ветровая эрозия, движение морского дна, вулканизм, горообразование и т. п.). Различают поэтому малые и большие круговороты в соответствии с объемом рассматриваемых экосистем: локальные биологические круговороты, биогеохимические циклы материков и океанов и, наконец, общепланетарный биогеохимический круговорот.
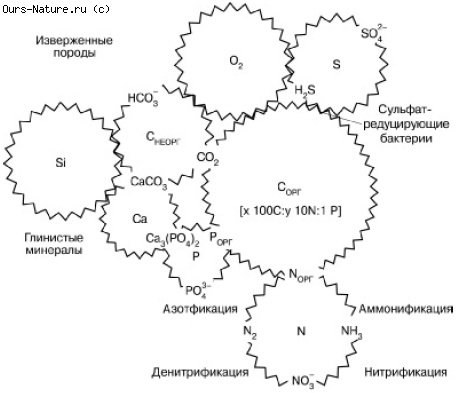
Рис. 163. Сопряжения циклов элементов в биосфере (по Г. А. Заварзину, 1998)
Вместе с тем отличительная черта биологических круговоротов – их неполная замкнутость. Часть химических элементов и их соединений постоянно выпадает из общей циркуляции и скапливается вне организмов, создавая своего рода запасы биогенных веществ. Так были накоплены кислород и азот в атмосфере, горючие ископаемые и другие породы земной коры. В масштабах геологического времени даже небольшое неравновесие в создании и разрушении органических веществ приводит к направленному преобразованию поверхностных слоев Земли.
Запасы биогенных веществ, мало доступные для живых организмов и медленно вовлекающиеся поэтому в биологический круговорот, составляют так называемый резервный фонд того или иного химического элемента в биосфере (например, кальция в карбонатных породах). Другая часть, интенсивно циркулирующая между телами организмов и окружающей средой, называется подвижным, или обменным, фондом. Соотношения между резервным и обменным фондами в биосфере у всех биогенных элементов различны.
Углерод. Углерод как химический элемент является основой жизни. Он может соединяться разными способами со многими другими элементами, образуя простые и сложные органические молекулы, входящие в состав живых клеток. По распространению на планете углерод занимает одиннадцатое место (0,35 % от веса земной коры), но в живом веществе в среднем составляет около 18 или 45 % от сухой биомассы.
В атмосфере углерод входит в состав углекислого газа СО2, в меньшей мере – в состав метана СН4 или следового количества других газообразных соединений. В гидросфере СО2 растворен в воде, и общее его содержание намного превышает атмосферное. Океан служит мощным буфером регуляции СО2 в атмосфере: при повышении в воздухе его концентрации увеличивается поглощение углекислого газа водой. Некоторая часть молекул СО2 реагирует с водой, образуя угольную кислоту, которая затем диссоциирует на ионы НСО3–и СО2‑3. Эти ионы реагируют с катионами кальция или магния с выпадением карбонатов в осадок. Подобные реакции лежат в основе буферной системы океана, поддерживающей постоянство рН воды. При подкислении (увеличении концентрации ионов Н+) происходит сдвиг влево в цепи: СО2 воздуха → СО2 воды → Н2СО3 → Са(НСО3)2 → СаСО3. При подщелачивании усиливается выпадение в осадок карбонатов кальция.
Углекислый газ атмосферы и гидросферы представляет собой обменный фонд в круговороте углерода, откуда его черпают наземные растения и водоросли (рис. 164). Фотосинтез лежит в основе всех биологических круговоротов на Земле. Высвобождение фиксированного углерода происходит в ходе дыхательной активности самих фотосинтезирующих организмов и всех гетеротрофов – бактерий, грибов, животных, включающихся в цепи питания за счет живого или мертвого органического вещества.
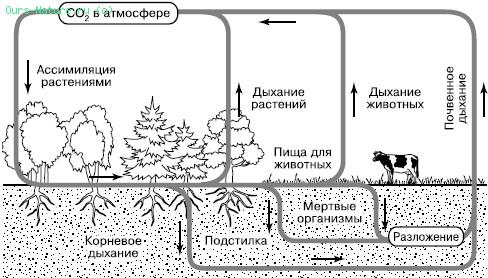
Рис. 164. Круговорот углерода в биосфере (по Б. Болину, 1972)
Особенно активно происходит возврат в атмосферу СО2 из почвы, где сосредоточена деятельность многочисленных групп деструкторов и редуцентов и осуществляется дыхание корневых систем растений. Этот интегральный процесс обозначается как «почвенное дыхание» и вносит существенный вклад в пополнение обменного фонда СО2 в воздухе. Параллельно с процессами минерализации органического вещества в почвах образуется гумус – богатый углеродом сложный и устойчивый молекулярный комплекс. Гумус является носителем почвенного плодородия, поскольку разрушается определенными группами микроорганизмов медленно и постепенно, обеспечивая равномерное питание растений. Гумус почв является одним из важных резервуаров углерода на суше.
В тех условиях, где деятельность деструкторов тормозится факторами внешней среды (например, при возникновении анаэробного режима в почвах и на дне водоемов), органическое вещество, накопленное растительностью, не разлагается, превращаясь со временем в такие породы, как каменный или бурый уголь, торф, сапропели, горючие сланцы и другие, богатые накопленной солнечной энергией. Они пополняют собой резервный фонд углерода, надолго выключаясь из биологического круговорота. Углерод временно депонируется также в живой биомассе, в мертвом опаде, в растворенном органическом веществе океана и т. п. Однако основным резервным фондом углерода на планете являются не живые организмы и не горючие ископаемые, а осадочные породы – известняки и доломиты. Их образование также связано с деятельностью живого вещества. Углерод этих карбонатов надолго захоранивается в недрах Земли и поступает в круговорот лишь в ходе эрозии при обнажении пород в тектонических циклах.
В биологическом круговороте участвуют лишь доли процента углерода от общего его количества на Земле. Углерод атмосферы и гидросферы многократно проходит через живые организмы. Растения суши способны исчерпать его запасы в воздухе за 4–5 лет, запасы в почвенном гумусе – за 300–400 лет. Основной возврат углерода в обменный фонд происходит за счет деятельности живых организмов, и лишь небольшая часть его (тысячные доли процента) компенсируется выделением из недр Земли в составе вулканических газов.
В настоящее время мощным фактором перевода углерода из резервного в обменный фонд биосферы становится добыча и сжигание огромных запасов горючих ископаемых.
По учетам в сети глобального мониторинга это уже приводит к повышению концентрации СО2 в атмосфере, последствия чего для судьбы человеческого общества усиленно обсуждаются и требуют научно обоснованного прогнозирования.
Кислород. С углеродным циклом теснейшим образом сопряжен круговорот кислорода.
Своей уникальной среди планет атмосферой с высоким содержанием свободного кислорода Земля обязана процессу фотосинтеза. Кислород освобождается из молекул воды и является по сути дела побочным продуктом фотосинтетической активности растений. Абиотическим путем кислород возникает в верхних слоях атмосферы за счет фотодиссоциации паров воды, но этот источник составляет лишь тысячные доли процента от поставляемыми фотосинтезом. Между содержанием кислорода в атмосфере и гидросфере существует подвижное равновесие. В воде его примерно в 21 раз меньше на равный объем по сравнению с воздухом.
Выделившийся кислород интенсивно расходуется на процессы дыхания всех аэробных организмов и на окисление разнообразных минеральных соединений. Эти процессы происходят в атмосфере, почве, воде, илах и горных породах. Показано, что значительная часть кислорода, связанного в осадочных породах, имеет фотосинтетическое происхождение. Обменный фонд О2 в атмосфере составляет не более 5 % от общей продукции фотосинтеза. Многие анаэробные бактерии также окисляют органические вещества в процессе анаэробного дыхания, используя для этого сульфаты или нитраты.
Накопление кислорода в атмосфере и гидросфере происходит в геологической истории в результате неполной замкнутости цикла углерода. На полное разложение органического вещества, создаваемого растениями, требуется точно такое же количество кислорода, которое выделилось при фотосинтезе. Захоронение органики в осадочных породах, углях, торфах послужило основой поддержания обменного фонда кислорода в атмосфере. Весь имеющийся в ней кислород проходит полный цикл через живые организмы примерно за 2000 лет.
В настоящее время значительная часть кислорода атмосферы связывается в результате работы транспорта, промышленности и других форм антропогенной деятельности. По расчетам, человечество тратит уже более 10 млрд т свободного кислорода из общего количества в 430–470 млрд т поставляемых процессами фотосинтеза. Если учесть, что в обменный фонд поступает лишь небольшая часть фотосинтетического кислорода, деятельность людей в этом отношении начинает приобретать угрожающие масштабы.
Вода. В ходе фотосинтеза растения используют водород воды в построении органических соединений, освобождая молекулярный кислород. В процессах дыхания всех живых существ, при окислении органических соединений вода образуется вновь. В истории жизни вся свободная вода гидросферы многократно прошла циклы разложения и новообразования в живом веществе планеты.
Кроме биологических циклов на Земле также осуществляется глобальный круговорот воды, движимый энергией Солнца. Вода испаряется с поверхности водоемов и суши и затем вновь поступает на Землю в виде осадков. Над океаном испарение превышает осадки, над сушей – наоборот. Эти различия компенсируются речным стоком. В глобальном круговороте воды растительность суши играет немаловажную роль. Транспирация растений на отдельных участках земной поверхности может составить до 80–90 % выпадающих здесь осадков, а в среднем по всем климатическим поясам – около 30 %. В истории биосферы появление наземной растительности вызвало за счет транспирации перераспределение осадков над морем и сушей и тем самым косвенно повлияло на размеры речного стока и процессы физического и химического выветривания.
Азот. В атмосфере и живом веществе содержится менее 2 % всего азота на Земле, но именно он поддерживает жизнь на планете. Азот входит в состав важнейших органических молекул – ДНК, белков, липопротеидов, АТФ, хлорофилла и других. В растительных тканях его соотношение с углеродом составляет в среднем 1: 30, а в морских водорослях 1: 6. Биологический цикл азота поэтому также тесно связан с углеродным.
Молекулярный азот атмосферы недоступен растениям, которые могут усваивать этот элемент только в виде ионов аммония, нитратов или из почвенных или водных растворов. Поэтому недостаток азота часто является фактором, лимитирующим первичную продукцию. Тем не менее атмосферный азот широко вовлекается в биологический круговорот благодаря деятельности прокариотических организмов (рис. 165). Способностью к фиксации молекулярного азота обладают очень многие прокариоты. В большой мере она развита у фотосинтезирующих сине‑зеленых водорослей (цианобактерий). Активно фиксируют азот свободно живущие в почве бактерии рода Azotobacter, а также клубеньковые бактерии Rhizobium, живущие на корнях растений семейства бобовых. При этом может связываться до 400 кг азота на га в год. Отмирая, бактериальные клетки обогащают почву азотными соединениями, доступными для растений. Симбиотические формы снабжают хозяина и за счет прижизненных выделений.
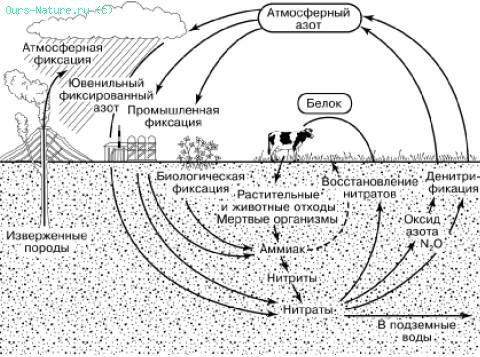
Рис. 165. Круговорот азота в биосфере (по К. Делвичу, 1972)
В круговороте азота принимают большое участие также аммонифицирующие микроорганизмы. Они разлагают белки и другие содержащие азот органические вещества до образования аммиака. В аммонийной форме азот частью вновь поглощается корнями растений, а частью перехватывается нитрифицирующими микроорганизмами. Они используют для себя химическую энергию, окисляя аммиак сначала в нитритную форму (бактерии Nitrosomonas). Окисление нитритов в нитраты производят бактерии рода Nitrobacter. Образовавшиеся нитраты вновь используются растениями в ходе фотосинтеза.
Прямо противоположна по функциям группа микроорганизмов – денитрификаторов.
В анаэробных условиях в почвах или водах они используют кислород нитратов для окисления органических веществ, получая энергию для своей жизнедеятельности. Азот при этом восстанавливается до молекулярного. Азотфиксация и денитрификация в природе приблизительно уравновешены. Цикл азота, таким образом, зависит преимущественно от деятельности бактерий, тогда как растения встраиваются в него, используя промежуточные продукты этого цикла и намного увеличивая масштабы азотной циркуляции в биосфере за счет продуцирования своей биомассы. Объемы микробной фиксации азота составляют до 2,5 т на км2 в год.
Небиологическая фиксация азота и поступление в почвы его окислов и аммиака происходит также с дождевыми осадками при ионизации атмосферы и грозовых разрядах. В среднем это дает около 1 т связанного азота на км2 в год.
Современная промышленность удобрений фиксирует азот атмосферы в размерах, превышающих природную азотфиксацию в целях увеличения продукции сельскохозяйственных растений.
Высокие дозы азотных удобрений приводят, однако, к вымыванию нитратов в грунтовые воды, водоемы и в конечном счете – питьевую воду, а также к избытку их в продуктах питания, что является опасным для человека. Таким же источником загрязнений служат сточные воды с высоким содержанием аммония. На его окисление до нитратов тратится растворенный в воде кислород, что часто бывает губительно для гидробионтов. Таким образом, деятельность человека все сильнее влияет на круговорот азота, в основном в сторону превышения перевода его в связанные формы над процессами возврата в молекулярное состояние.
Фосфор. Этот элемент, необходимый для синтеза многих органических веществ, включая АТФ, ДНК, РНК, усваивается растениями только в виде ионов ортофосфорной кислоты (РО3+4). Он относится к элементам, лимитирующим первичную продукцию и на суше, и особенно в океане, поскольку обменный фонд фосфора в почвах и водах невелик. Круговорот этого элемента в масштабах биосферы незамкнут.
На суше растения черпают из почвы фосфаты, освобожденные редуцентами из разлагающихся органических остатков, но в щелочной или кислой почве растворимость фосфорных соединений резко падает. Основной резервный фонд фосфатов содержится в горных породах, созданных на дне океана в геологическом прошлом. В ходе выщелачивания пород часть этих запасов переходит в почву и в виде взвесей и растворов вымывается в водоемы. В гидросфере фосфаты используются фитопланктоном, переходя по цепям питания в других гидробионтов. Однако в океане большая часть фосфорных соединений захоранивается с остатками животных и растений на больших глубинах и не попадает вновь в фотическую зону, переходя с осадочными породами в большой геологический круговорот. На глубине растворенные фосфаты связываются с кальцием, образуя фосфориты и аппатиты. В биосфере, по сути дела, происходит однонаправленный поток фосфора из горных пород суши в глубины океана, и обменный фонд его в гидросфере очень ограничен. Биологическая продуктивность океана резко повышается в районах, где увеличивается доступность фосфора для фитопланктона – при сносе его речными водами, у побережий морей и в зонах так называемого апвеллинга – восходящих океанических течений, возвращающих фосфорные соединения со дна в освещенные слои воды.
Наземные залежи фосфоритов и аппатитов разрабатываются в качестве удобрений. Снос избыточного фосфора в пресные водоемы является одной из главных причин их «цветения» – бурной эвтрофикации.
Сера. Круговорот серы, необходимой для построения ряда аминокислот, ответственных за трехмерную структуру белков, поддерживается в биосфере широким спектром бактерий. В отдельных звеньях этого цикла участвуют аэробные микроорганизмы, окисляющие серу органических остатков до сульфатов, а также анаэробные сульфатредукторы, восстанавливающие сульфаты до сероводорода, кроме них – разные группы серобактерий, окисляющих сероводород до элементарной серы и далее – сульфатов, тионовые бактерии, переводящие элементарную серу также в соли серной кислоты. Растения усваивают из почвы и воды только ионы SO2‑4, поставляемые им деятельностью прокариотов.
Основное накопление серы происходит в океане, куда сульфатные ионы непрерывно поступают с суши с речным стоком. Частично сера возвращается в атмосферу при выделении из вод сероводорода и окисляется здесь до двуокиси, превращаясь в дождевой воде в серную кислоту. Промышленное использование большого количества сульфатов и элементарной серы и сжигание горючих ископаемых поставляют в атмосферу большие объемы диоксида серы. Это вредит растительности, животным, людям и служит источником кислотных дождей, усугубляющих отрицательные эффекты вмешательства человека в круговорот серы.
Как мы видим, круговороты всех биогенных элементов на планете поддерживаются сложным взаимодействием разных частей биосферы. Они формируются деятельностью разных по функциям групп организмов, системой стока и испарения, связывающих океан и сушу, процессами циркуляции вод и воздушных масс, действием сил гравитации, тектоникой плит и другими масштабными геологическими и геофизическими процессами. Биосфера действует как единая сложная система, в которой циркулируют с разной скоростью атомы отдельных элементов. Однако главным двигателем этих круговоротов является живое вещество планеты, все живые организмы, обеспечивающие процессы синтеза, трансформации и разложения органического вещества.
10.2. Геохимическая работа живого вещества10.4. Развитие биосферы