Книга: Новая наука о жизни
Навигация: Начало Оглавление Другие книги
2.4. Органицизм
Организмические теории морфогенеза развивались под различными влияниями: некоторые – из философских систем, особенно теории А. Н. Уайтхеда и Дж. К. Сматса; другие – из современной физики, в частности из теории поля; третьи – из гештальт‑психологии, которая сама испытывала сильное воздействие концепции физических полей; а некоторые – из витализма Дриша.[77]
Эти теории рассматривают те же проблемы, которые Дриш считал неразрешимыми с механистических позиций, – регуляцию, регенерацию и репродукцию, – но там, где для объяснения свойств целостности и направленности, проявляемых развивающимися организмами, Дриш предлагал нефизическую энтелехию, органицисты предложили морфогенетические (или эмбриональные, или свойственные развитию) поля.
Эта идея была выдвинута независимо А. Гурвичем в 1922[78] и П. Вейсом в 1926 году.[79] Однако, помимо утверждения, что морфогенетические поля играют важную роль в контроле морфогенеза, ни один из этих авторов не определял, что такое эти поля и как они работают. Терминология полей вскоре была воспринята другими учеными, работающими в области биологии развития, но она оставалась слабо разработанной, хотя и служила для создания аналогий между свойствами живых организмов и неорганических электромагнитных систем. Например, если магнит из железа разрезан на две половинки, получаются два целых магнита, благодаря свойствам магнитного поля; подобно этому, морфогенетические поля предлагались как объяснение «целостности» отделенных частей организмов, которые были способны вырасти в целые организмы.
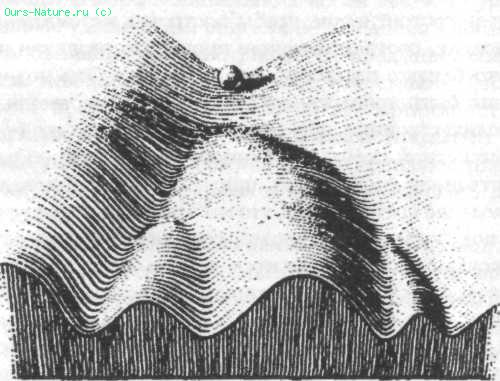
Рис. 5. Часть «эпигенетического ландшафта», иллюстрирующая концепцию хреоды как канализированного пути изменения (из Waddington, 1957)
К. X. Уоддингтон предложил более широкое толкование морфогенетических полей, включив в рассмотрение временной аспект развития. Он назвал эту новую концепцию хреодой (от греческого chrē – необходимо и hodos – путь) и иллюстрировал ее с помощью простого трехмерного «эпигенетического ландшафта» (рис. 5) .[80] В этой модели путь, по которому движется шарик, когда он катится вниз, соответствует истории развития определенной части яйца. По мере развития эмбриологии появляются разветвляющиеся серии альтернативных путей развития, представленные «долинами». Они соответствуют путям развития различных типов органа, ткани и клетки. В организме они вполне различимы: например, почка и печень имеют различные структуры и не переходят друг в друга через серию промежуточных форм. Развитие канализировано по направлению к определенным конечным точкам. Генетические изменения, или пертурбации, в окружающей среде могут «толкнуть» направление развития (представленное путем, по которому движется шарик) из глубины «долины» на соседний «холм», но, если оно не будет переброшено через вершину холма в соседнюю долину, процесс развития найдет путь назад. Он вернется не к начальной точке, а к более поздней позиции на канализованном пути изменений. Это представляет регуляцию.
Концепция хреоды очень близка идее морфогенетических полей, но она делает явным измерение времени, которое в морфогенетических полях присутствует в неявном виде.
Недавно обе эти концепции были существенно продвинуты математиком Рене Томом, который сделал попытку создать математическую теорию, описывающую не только морфогенез, но также поведение и язык.[81] Его главной задачей было найти подходящий математический формализм для этих проблем, которые до сих пор не поддавались математической обработке. Конечная цель состоит в том, чтобы построить математические модели, соответствующие процессам развития настолько близко, насколько это возможно. Эти модели должны быть топологические, скорее, качественные, чем количественные, и не должны зависеть от какой‑либо конкретной схемы причинного объяснения: «Одна существенная особенность применения нами локальных моделей состоит в том, что мы ничего не подразумеваем под "конечной природой реальности"; даже если она когда‑либо обнаружится посредством анализа, слишком сложного для описания, только часть ее проявления – так называемые «наблюдаемые» (observ‑ables) в конечном счете пригодны для макроскопического описания системы. Фазовое пространство нашей динамической модели определено с использованием лишь таких «наблюдаемых» и без ссылок на какие‑либо более или менее хаотические базовые структуры».[82]
Проблема с этим подходом состоит в том, что он является по существу описательным; он мало что дает для объяснения морфогенеза. Это характерно для всех существующих организмических теорий морфогенеза. Сравним, например, хреоду Уоддингтона и энтелехию Дриша. Обе заключают в себе идею о том, что развитие направляется и располагается (канализируется) в пространстве и времени чем‑то, что само не может рассматриваться как принадлежащее определенному месту и времени; оба видят это что‑то как содержащее в самом себе конец и цель процесса развития, и вследствие этого оба предлагают способ объяснения регуляции. Главное различие между ними состоит в том, что Дриш пытался разъяснить, как предложенный им процесс может работать на практике, а Уоддингтон этого не делал. Концепция хреоды была менее уязвима для критики, потому что оставалась столь неопределенной.[83] Фактически, Уоддингтон рассматривал концепции хреоды и морфогенетических полей как «по существу способ, удобный для описания».[84] Подобно многим другим органицистам, он отрицал, что предлагает оперировать чем‑либо другим, нежели известными физическими причинами.[85] Однако не все органицисты это отрицали, некоторые оставляли вопрос открытым. Такое явно уклончивое поведение можно проиллюстрировать следующим обсуждением морфогенетического поля у Б. К. Гудвина:
«Один аспект поля состоит в том, что на него могут влиять электрические силы. Было обнаружено, что другие развивающиеся и регенерирующие организмы имеют интересную и значительную электрическую сеть, но я не хотел бы предположить, что морфогенетическое поле имеет по существу электрическую природу. Химические вещества также влияют на полярность и другие пространственные аспекты развивающихся организмов; но опять‑таки я не хотел бы делать отсюда вывод, что морфогенетическое поле имеет по существу химическую или биохимическую природу. Мое убеждение состоит в том, что исследование этого поля должно проводиться при допущении, что оно имеет природу какую‑либо из упомянутых, или никакую из них, или все сразу; но я считаю, что, несмотря на агностицизм в отношении его материальной природы, оно играет главную роль в процессе развития» .[86]
Открытость этой позиции делает ее наиболее многообещающей отправной точкой для построения обстоятельной организмической теории морфогенеза. Но очевидно, что, если морфогенетические поля считаются полностью объяснимыми с помощью известных физических принципов, они представляют собой не что иное, как неясную терминологию, наложенную на усложненную версию механистической теории. Только если допускается, что они играют причинную роль, не признанную физикой сегодня, может быть построена теория, доступная проверке. Такая возможность исследуется в следующих главах.
2.3. ВитализмГлава 3. Причины формы