Книга: Новая наука о жизни
Навигация: Начало Оглавление Другие книги
8.3. Дивергенция хреод
Если мутация или изменение окружения нарушает нормальный ход морфогенеза на сравнительно ранней стадии, система может быть способна к регулированию и продолжению развития до создания нормальной формы, несмотря на это нарушение. Если этот процесс повторяется поколение за поколением, отклонение хреоды будет стабилизироваться морфическим резонансом; следовательно, вся порода или разновидность данного вида начнет следовать ненормальному пути морфогенеза, хотя в результате получится обычная взрослая форма. Действительно, описано много случаев так называемых временных отклонений в развитии. Например, у торфяных червей Prorhynchnus stagnitilis яйцеклетки делятся либо спиральным, либо радиальным способом, и развивающиеся эмбрионы растут либо внутри желтка, либо на его поверхности. Вследствие этих различий на ранних стадиях эмбрионального развития некоторые органы формируются в различной последовательности; тем не менее взрослые животные идентичны. А у единственного вида кольчатого червя, Nereis, образуются два очень разных типа личинок; несмотря на это, обе развиваются в одну и ту же взрослую форму.[170] В некоторых из таких случаев временные отклонения могут быть адаптивными, например, к условиям жизни личинок, но по большей части они возникают без видимых причин. Гораздо большее эволюционное значение имеют те отклонения хреод, которые не полностью корректируются путем регуляции и поэтому приводят к появлению отличающихся друг от друга конечных форм. Такие изменения путей развития могут возникать в результате мутаций (ср. раздел 7.3) или необычных условий окружающей среды (ср. раздел 7.6). В случае мутации в неизменном окружении, если отклонившаяся конечная форма имеет преимущество с точки зрения отбора, частота мутантных генов в популяции будет возрастать и, кроме того, новая хреода усилится вследствие морфического резонанса. В более сложном случае, когда отклонившаяся форма, возникшая в ответ на необычные окружающие условия, имеет селективное преимущество, новая хреода будет усиливаться, как и раньше, и в то же время отбор будет действовать в пользу организмов, имеющих генетические возможности для такой реакции на окружение (сравните с экспериментами Уоддингтона на плодовых мушках, раздел 7.7). Так приобретенные признаки станут наследуемыми в результате сочетания генетического отбора и морфического резонанса.
В естественных условиях действие различных давлений отбора на географически или экологически изолированные популяции какого‑либо вида приведет к расхождению (дивергенции) как их «генных пулов», так и их хреод. Действительно, бесчисленные виды животных и растений разошлись (в ходе эволюции) в генетически и морфологически различные породы и разновидности; знакомыми примерами являются одомашненные животные и культурные растения.[171] Подумайте, например, о поразительно различающихся породах собак, от афганской гончей до китайского мопса.
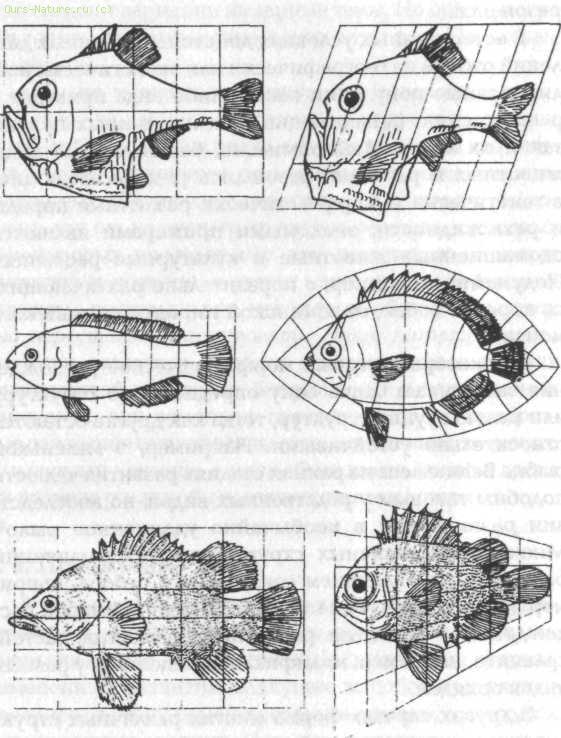
Рис. 20. Сравнение форм различных видов рыб (из Thompson, 1942). Воспроизведено с разрешения издательства Cambridge University Press
В некоторых случаях морфологическое расхождение затрагивает лишь одну определенную структуру или малую группу структур, тогда как другие остаются относительно устойчивыми. Например, в маленькой рыбке Beloneacus на ранних стадиях развития челюсти подобны таковым у родственных видов, но впоследствии развиваются в необычайно удлиненное рыло.[172] Множество различных структурных преувеличений развились под влиянием сексуального отбора, например рога у оленя. А цветы дают тысячи примеров расхождений в развитии различных составных частей: сравните, например, модификации лепестков у разных видов орхидей.
В других случаях форма многих различных структур меняется сходным образом. Если формы варьируются особенно единообразно и гармонично, их можно сравнивать графически, по систематическому искривлению наложенных координат (рис. 20), как показал сэр Д'Арси Томпсон в разделе своего очерка «О росте и форме», озаглавленном «Теория трансформаций, или сравнение родственных форм».
Эволюционные изменения такого типа происходят в рамках уже существующих морфогенетических полей. Они создают вариации на данные темы. Но они не могут объяснить существование самих этих тем. По словам Томпсона:
«Мы не можем превратить беспозвоночное в позвоночное или кишечнополостное в червя посредством какой‑либо простой и правильной деформации или чего‑либо, не поддающегося разложению на элементарные принципы… Формальное сходство, от которого мы зависим как от надежного критерия родства животных в некоторых пределах или при некоторых степенях близости и подобия, в других случаях перестает нам служить, поскольку при некоторых обстоятельствах оно перестает существовать. Наши геометрические аналогии являются весомым свидетельством против дарвиновской концепции бесчисленных и непрерывных изменений; они помогают показать, что скачкообразные изменения естественны, что. внезапные изменения, большие или меньшие, должны были иметь место и время от времени должны были возникать новые типы» .[173]
8.2. Мутации8.4. Подавление хреод