Книга: Путешествие в страну микробов
Навигация: Начало Оглавление Другие книги
Генетический алфавит
Авторы модели ДНК представляют систему «кодирования» следующим образом: различные свойства ДНК в разных организмах объясняются различием в составе нуклеотидов в ее молекуле. Нуклеотиды, по их мнению, являются своего рода генетическим алфавитом, при помощи которого в ДНК «химическим почерком» закодированы наследственные особенности организма. А поскольку ДНК содержит по меньшей мере несколько тысяч нуклеотидов, возникают неограниченные возможности комбинаций во взаимном чередовании и порядке пар А — Т, Г — Ц в ее молекуле. Каждая новая комбинация придает новые свойства ДНК.
Молекулы ДНК в клетке, поясняют далее Дж. Уотсон и Ф. Крик, способны к «самовоспроизведению» (редупликации) с сохранением постоянного расположения нуклеотидов. Мы знаем, что внутренний «цилиндр» молекулы ДНК состоит из пар оснований А — Т и Г — Ц, соединенных между собой водородными связями. Редупликация молекул ДНК может идти следующим образом. Водородные связи между парными нуклеотидами на одном конце молекулы постепенно начинают прерываться и обе спиральные цепи ДНК «разматываются», освобождаясь друг от друга. Такая развернувшаяся цепь удерживается связями между остатками молекул дезоксирибозы и фосфорной кислоты, «выставив» перпендикулярно оси свои основания. В окружающей среде находятся синтезированные клеткой свободные нуклеотиды, которые могут входить в реакцию со свободными основаниями развернутой цепи ДНК. Но к каждому основанию может приблизиться и соединиться с ним только нуклеотид, имеющий парное, «дополняющее» основание. Это значит, что к развернутой цепи начнет присоединяться другая, недостающая цепь ДНК, причем точно такая, какая была в другой половине макромолекулы и отделилась, чтобы подобным же образом извлечь из среды то, чего ей недостает до полной молекулы ДНК. В результате этих процессов образуются две молекулы ДНК, каждая из которых имеет половину материнской молекулы, дополненную вновь синтезированной. Дочерние молекулы становятся, таким образом, точной копией материнской ДНК. При этом сохраняется и состав генетического материала.
Здесь мы должны, однако, предупредить читателя, что то, о чем он только что прочел, было всего-навсего гипотезой Уотсона и Крика о редупликации ДНК. Вы можете справедливо заметить, что правильность гипотез должна быть доказана экспериментально.
Описанную гипотезу подтвердил американский биохимик А. Корнберг, которому удалось открыть полимеразу ДНК — фермент, участвующий в процессе редупликации. Получив из клеток кишечных бактерий Escherichia coli ДНК и фермент, он соединил их в пробирке, добавив туда же смесь соответствующих нуклеотидов. Через некоторое время количество ДНК в пробирке значительно увеличилось, причем были использованы присутствующие в среде свободные Нуклеотиды. За свое открытие он получил в 1959 году Нобелевскую премию по физиологии и медицине. Эту премию разделил с ним профессор С. Очоа, открывший независимо от Корнберга фермент — полимеразу РНК, которая синтезирует рибонуклеиновую кислоту. Совсем недавно Корнберг обнаружил еще один фермент, участвующий в синтезе ДНК, и назвал его фосфатазой ДНК.
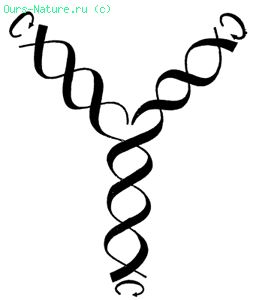
Модель редупликации молекулы ДНК. К развернутым цепочкам (вверху) присоединяются дополнительные цепочки того же состава, что и в материнской молекуле.
Гипотезу о редупликации ДНК подтвердили в 1958 году М. С. Месельсон и Ф. Сталь. Они культивировали бактерии Е. coli в жидкой питательной среде, содержащей вещества с радиоактивным азотом 15N. ДНК этих бактерий оказалась потом «меченной» радиоактивным элементом всюду, где в ее макромолекулах содержится атом азота. Затем ученые культивировали бактерии в жидкой питательной среде, содержащей нерадиоактивный азот 14N. При выращивании бактерии размножались, и можно было наблюдать, как радиоактивный азот ДНК равномерно распределялся по дочерним молекулам ДНК. В каждой последующей генерации бактерий отмечалась половинная радиоактивность ДНК по сравнению с предыдущей. Иначе говоря, при каждой редупликации ДНК радиоактивность материнской макромолекулы равномерно распределялась в двух дочерних макромолекулах, которые создавали недостающие им цепи, привлекая для этого нуклеотиды с нормальным азотом.
Днк в роли диктатораЖакоб и моно обдумывают генетический код