Книга: Волк. Вопросы онтогенеза поведения, проблемы и метод реинтродукции
Навигация: Начало Оглавление Другие книги
Полученные результаты и их обсуждение
Прежде чем изложить материалы, полученные в результате наблюдения за животными, целесообразно привести таблицу коэффициентов, отражающих уровень браконьерства в изученных нами регионах (таблица 16).
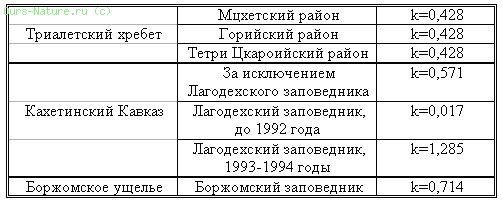
Таблица 16.
Коэффициент уровня браконьерства в различных регионах Грузии.
Изучение дистанции бегства от нас диких копытных показало, что ее показатель находится в прямой зависимости от коэффициента (к) уровня браконьерства. Так, например, дистанция бегства от нас была минимальной в тех районах, где к=0,017 (кавказский олень – 30–50 м), а там, где коэффициент браконьерства был максимальным (к=1,285) соответственно увеличивалась и дистанция бегства (кавказский олень – 300–500). Детально эти данные приведены в таблице 17.
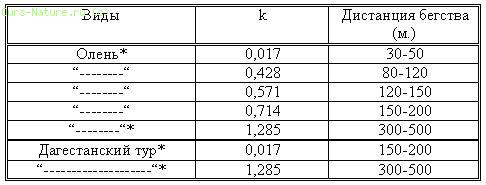
Таблица 17.
Влияние уровня браконьерства на дистанцию бегства копытных от человека. * – одни и те же популяции.
Данные об изменении уровня браконьерства в одном и том же регионе получены нами только в Лагодехском заповеднике. Однако и их достаточно, чтобы было очевидным влияние этого фактора на дистанцию бегства представителей одной и той же популяции (рис. 17). Полученный материал однозначно указывает на то, что дистанция бегства не является врожденной. Она может изменяться пропорционально изменению пресса браконьерства на копытных. Исходя из этого, об уровне браконьерства можно судить по дистанции бегства копытных от человека. По нашему мнению, это имеет большое практическое значение для оперативного выявления ситуации в том или ином районе.

Рисунок 17.
Зависимость дистанции бегства Кавказского благородного оленя от уровня браконьерства.
X – коэффициент браконьерства; Y – средняя дистанция бегства. * – одни и те же популяции.
Следует отметить тот факт, что дистанция бегства существующих в группе молодых индивидов и взрослых животных одинакова. Мы подчеркиваем, существующих в группе, т. к. дистанция бегства молодых может измениться после того, как они выйдут из‑под опеки родителей и начнут самостоятельную жизнь.
Полученный нами материал говорит и о том, что дистанция бегства взрослых индивидов меняется в зависимости от индивидуального (или группового) опыта. Потомству же опыт передается путем опосредованного обучения. Исходя из этого, следует подчеркнуть, что дистанция бегства индивида не является врожденной и передается следующему поколению за счет врожденной способности к подражанию. Последнее же – один из значительных механизмов обучения.
Эксперименты, проведенные при подкрадывании к диким копытным на возможно минимальную дистанцию, показали, что самки оленей и туров с детенышами при внезапном появлении человека на близком расстоянии издают специфические звуки (олени – резкий лающий звук, туры – свист). Вслед за этим убегают взрослые особи; детеныши же, как правило, убегают сразу при звуке тревоги. В некоторых случаях (n=27) детеныши убегали только вместе с родителями. При повторном подкрадывании к этим индивидам детеныши убегали не вместе с родителями, а на фоне звуков тревоги, издаваемых родителями. Описанные факты указывают на то, что реакция убегания детенышей формируется за счет одноразового обучения. Для этой формы обучения решающее значение имеет врожденная реакция на издаваемый родителем звук тревоги и сопряженная с этим звуком поведенческая реакция того же родителя. Следует подчеркнуть и то, что этот звук является для детеныша безусловным раздражителем. Именно сопряжение этого раздражителя с появлением человека обуславливает формирование защитного поведения детенышей за счет одноразового обучения. При сопоставлении материала, полученного нами на волках и копытных, стало очевидным, что и у хищных, и у копытных в процессе постнатального онтогенеза описанная поведенческая реакция должна формироваться по одному и тому же принципу. Можно предположить, что поведенческая реакция родителей формирует традицию конкретной формы защитного поведения потомства и ее передачу от поколения к поколению путем опосредованного обучения.
На основе вышесказанного можно сделать вывод, что ситуационное выражение и интенсивность защитной формы в зависимости от поведения не является врожденной и поэтому может меняться в зависимости от состояния окружающей среды. Этим обеспечивается формирование оптимальной формы поведения и ее интенсивности, и тем самым адаптация к условиям окружающей среды.
Наблюдения за охотой подготовленных нами волков показали, что с момента реинтродукции в течение 6 месяцев дистанция бегства от них диких копытных возросла приблизительно в 6 раз. Изменение дистанции бегства показано на графике 18.

Рисунок 18.
Изменение дистанции бегства косуль от реинтродуцированных волков.
X – месяцы со дня реинтродукции; Y – дистанция бегства в метрах.
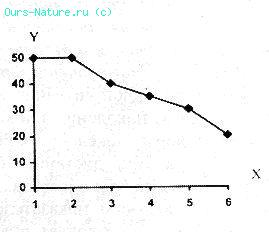
Рисунок 19.
Изменение процента успешной охоты реинтродуцированных волков.
X – месяцы со дня реинтродукции; Y – процент успешной охоты.
Как видно из приведенного материала, между дистанцией бегства от хищника и успешностью охоты последнего существует отрицательная корреляция (рис. 19).
Для формирования такой корреляции, по‑видимому, большое значение имеет опыт, приобретенный «спасшейся» жертвой. Этот опыт использует не только она сама, но и передает его другим (включая потомство) путем опосредованного обучения. Интересно и то, что дистанция бегства копытных увеличивается и в том случае, когда индивиды наблюдают смерть своих собратьев. Поэтому обучение возможно не только за счет собственного опыта или при наблюдении за завершенной поведенческой реакцией других особей, но и тогда, когда другой индивид успевает осуществить лишь небольшой фрагмент защитного поведения и оно кончается неудачей. Исходя из этого, одноразовое сочетание внешних признаков врага (хищник, человек) и защитного поведения или его фрагмента у собрата достаточно для формирования адекватной защитной реакции животного – «наблюдателя».
Сразу же после восстановления у копытных защитной реакции по отношению к волку в вышеуказанных регионах провели эксперименты с целью изучения их реакции на хищников, не имеющих опыта охоты. Эксперименты показали, что во всех 20 случаях у копытных развивалась такая же четко выраженная защитная реакция, как и на особей, имеющих опыт охоты. Это еще раз указывает на решающее значение видовых признаков хищника для формирования у копытных оборонительной реакции.
Эксперименты, проведенные в регионах постоянного обитания волка, показали, что реакция диких копытных на волков четко дифференцирована. В частности, на имеющих опыт охоты хищников у копытных имеется ярко выраженная защитная реакция, тогда как они индифферентны по отношению к тем особям, которые не умеют охотится, т. е. не воспринимают копытных как потенциальную жертву. Такие индивиды могли находиться в непосредственной близости к оленям. Приведенный материал перекликается с работой Мичурина (1970), где он отмечает, что часто видел волков, которые разгуливали среди группы оленей. Расстояние между ними менялось от 30 до 100 метров. В другом случае волк сидел среди большой группы оленей на холме. Приведенный нами материал указывает на то, что при взаимодействии с хищником копытные приобретают опыт, который позволяет им различать не только его видоспецифичные черты, но и специфику поведения этого вида и, исходя из этого, и его «цель».
Из всего вышесказанного можно сделать следующий вывод: реакция диких копытных на человека и хищников не является врожденной. Форма этой реакции и ее интенсивность могут изменяться соответственно уровню пресса на копытных. Приобретение опыта (обучение), может происходить с одного раза, не только во время непосредственно полученного «урока», но и методом наблюдения за реакцией других индивидов (опосредованное обучение). Для опосредованного обучения достаточно восприятия фрагмента реакции другого индивида, что полностью исключает необходимость приобретения собственного опыта, тем самым, значительно ускоряет процесс обучения и делает его значительно безопасным и, следовательно, имеет большое значение для выживания индивида.
В случае стабильной ситуации в ареале, опыт, приобретенный индивидом, передается из поколения в поколение путем опосредованного обучения и приобретает форму поведенческой традиции.
Как было показано, дистанция бегства копытных меняется в зависимости от уровня пресса браконьерства. Следовательно, сама дистанция бегства является безошибочным показателем уровня браконьерства. Наличие подобной зависимости позволяет оперативно определить ситуацию, существующую в том или ином регионе, что имеет важной практическое значение для контроля за уровнем браконьерства.
Если интенсивность описанной защитной реакции не является врожденной, то она может угаснуть или ослабнуть в том случае, если в ареале копытного, по разным причинам, больше нет хищника. Наглядным примером этого служит дистанция бегства косули от реинтродуцированных нами волков. Как оказалось, эта поведенческая форма и ее интенсивность восстановимы, но в масштабе популяции для ее восстановления необходимо длительное время. Исходя из этого, в процессе восстановления вида хищника очень важно, какое количество животных реинтродуцируется одновременно. Реинтродукция одновременно большого числа хищных до восстановления защитного поведения копытных может отрицательно сказаться на их популяцию.
Анализ приведенного материала позволяет сделать следующие выводы:
1. форма и интенсивность защитной реакции копытных по отношению к врагу (человеку или хищнику) подвержены сильной модификации в онтогенезе, вследствие чего возможны их изменения во времени в соответствии с существующей ситуацией;
2. в конкретной ситуации форма и интенсивность защитного поведения по отношению к врагу передается опосредованным путем из поколения в поколение;
3. у копытных, в случае отсутствия в их ареале хищников, через одно поколение гаснет оборонительная реакция по отношению к ним;
4. во избежание отрицательного воздействия на численность копытных, реинтродукция (судя по всему, и транслокация) восстанавливаемого вида хищников должна производиться небольшими группами и со значительными интервалами. Естественно, что интервал должен определяться, исходя из поведенческой и репродуктивной специфики копытных;
5. пресс браконьерства по отношению к копытным можно определять по дистанции их бегства от человека.
Материал и методикаЗАКЛЮЧЕНИЕ