Книга: Общая экология
Навигация: Начало Оглавление Другие книги
3.3.1. Адаптация растений к поддержанию водного баланса
Низшие наземные растения из влажного субстрата поглощают воду погруженными в него частями таллома, а влагу дождя, росы и тумана – всей поверхностью. В максимально набухшем состоянии лишайники содержат в 20–30 раз больше воды, чем сухого вещества.
Среди высших наземных растений мохообразные поглощают воду из почвы ризоидами, а большинство других – корнями, специализированными органами, всасывающими воду. В клетках корня развивается сосущая сила чаще всего в несколько атмосфер, но этого достаточно для извлечения из почвы большей части связанной воды. Лесные деревья умеренной зоны развивают сосущую силу корней около 3 · 106 Па (30 атм), некоторые травянистые растения (земляника лесная, медуница неясная) – до 2 · 106 (20 атм) и даже свыше 4 · 106 Па (40 атм) (смолка обыкновенная); растения сухих областей – до 60 атм.
Когда в непосредственной близости от корней запасы воды в почве истощаются, корни растут в направлении большей влажности, так что корневая система растений постоянно находится в движении. У степных и пустынных растений часто можно видеть эфемерные корни, быстро вырастающие в периоды увлажнения почвы, а с наступлением засушливого периода засыхающие.
По типу ветвления различают следующие корневые системы:
1) экстенсивная охватывает большой объем почвы, но сравнительно слабо ветвится, так что почва пронизана корнями негусто. Таковы корневые системы у многих степных и пустынных растений (саксаула, верблюжьей колючки), у деревьев умеренной полосы (сосны обыкновенной, березы повислой), а из трав у люцерны серповидной, василька шероховатого и др.;
2) интенсивная – охватывает сравнительно небольшой объем почвы, но густо пронизывает ее многочисленными сильно ветвящимися корнями, как, например, у степных дерновинных злаков (ковылей, типчака и др.), у ржи, пшеницы. Между этими типами корневых систем есть переходные.
Корневые системы очень пластичны и резко реагируют на изменение условий, в первую очередь увлажнения (рис. 29). При недостатке влаги корневая система становится экстенсивнее. Так, при выращивании ржи в разных условиях общая длина корней (без корневых волосков) в 1000 см3 почвы варьирует от 90 м до 13 км, а поверхность корневых волосков может увеличиться в 400 раз.
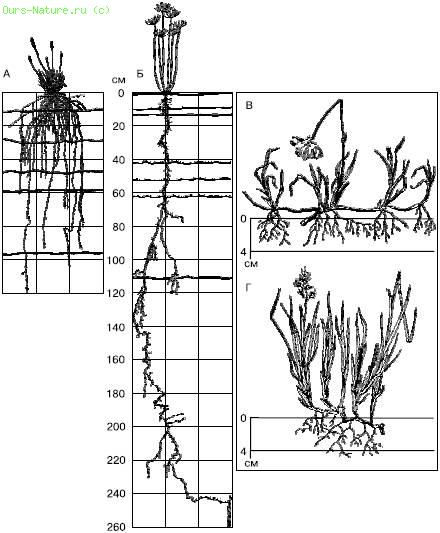
Рис. 29. Корневые системы стенных и тундровых растений (по М. С. Шалыту и Б. А. Тихомирову, 1963):
А – Festuca sulcata;
Б – Euphorbia gerardiana на черноземах в Аскания‑Нова;
В – Eriophorum scheuchzeri;
Г – Hierochloe alpina – из тундр Таймыра
Всасывание воды корнями затруднено при большой сухости почвы, засолении или сильной кислотности, при низкой температуре. Например, ясень обыкновенный при температуре почвы 0 °C поглощает воды в 3 раза меньше, чем при +(20–30) °С. Способность поглощать воду при той или иной температуре зависит от приспособленности растений к тепловому режиму почв в местах их произрастания. Виды с ранним началом развития, как правило, могут всасывать воду корнями при более низкой температуре, чем развивающиеся позднее. Тундровые растения и некоторые деревья, растущие на почвах с подстилающей их многолетней мерзлотой, могут поглощать воду при температуре почвы 0 °C.
У высших растений есть и дополнительные пути поступления воды в тело. Мхи могут поглощать воду всей поверхностью, как и лишайники. Особенно много воды впитывают такие мхи, как кукушкин лен, виды сфагнума, чему способствует строение их листьев и побегов. При полном насыщении сфагновые мхи содержат в своем теле в десятки раз больше воды, чем в воздушно‑сухом состоянии (рис. 30). Семена поглощают воду из почвы. Из воздуха, насыщенного водяными парами, в дождевом тропическом лесу поглощают воду многие эпифиты, например папоротник гименофиллум – тонкими листьями, многие орхидеи – воздушными корнями. В чашевидных влагалищах листьев многих зонтичных скапливается вода, которая постепенно всасывается эпидермисом. Виды из рода тилляндсия (бромелиевые) существуют в пустыне Атакама практически исключительно за счет влаги туманов и росы, которую впитывают чешуевидные волоски на листьях.
Поступившая в растение вода транспортируется от клетки к клетке (ближний транспорт) и по ксилеме во все органы, где расходуется на жизненные процессы (дальний транспорт). В среднем 0,5 % воды идет на фотосинтез, а остальная – на восполнение потерь от испарения и поддержание тургора. Вода испаряется со всех поверхностей, как внутренних, так и наружных, соприкасающихся с воздухом. Различают устьичную, кутикулярную и перидермальную транспирацию.
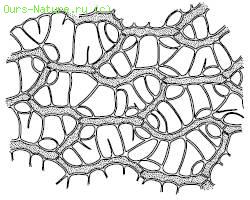
Рис. 30. Часть листа сфагнума болотного с клетками двух типов: зеленых с хлоропластами и бесцветных водоносных (Жизнь растений. Т. 4. 1978)
Через устьица транспирируется влага, испарившаяся с поверхности клеток внутри органов. Это основной путь расходования воды растением. Кутикулярная транспирация составляет менее 10 % от свободного испарения; у вечнозеленых хвойных пород она сокращается до 0,5 %, а у кактусов даже до 0,05 %. Относительно велика кутикулярная транспирация молодых развертывающихся листьев. Перидермальная транспирация обычно незначительна. Интенсивность общей транспирации повышается с увеличением освещенности, температуры, сухости воздуха и при ветре.
Водный баланс остается уравновешенным в том случае, если поглощение воды, ее проведение и расходование гармонично согласованы друг с другом. Нарушения его могут быть кратковременными или длительными. По приспособлениям наземных растений к кратковременным колебаниям условий водоснабжения и испарения различают пойкилогидрические и гомойогидрические виды.
У пойкилогидрических растений содержание воды в тканях непостоянно и сильно зависит от степени увлажнения окружающей среды. Они не могут регулировать транспирацию и легко и быстро теряют и поглощают воду, используя влагу росы, туманов, кратковременных дождей, в сухом состоянии находятся в анабиозе. Способны обитать там, где короткие периоды увлажнения чередуются с длительными периодами сухости.
Пойкилогидричность свойственна цианобактериям, всем водорослям, некоторым грибам, лишайникам (рис. 31), а также ряду высших растений: многим мхам, некоторым папоротникам и даже отдельным цветковым, по‑видимому, вторично перешедшим к пойкилогидрическому образу жизни. Таков, например, южно‑африканский кустарник Myrothamnus flabel‑lifolia (розоцветные).

Рис. 31. Лишайник сосновых лесов Cetraria islandica – типичный пойкилогидрический организм (Жизнь растений. Т. 3. 1978)
В мелких клетках таллома большинства низших растений нет центральной вакуоли, поэтому при высыхании они равномерно сжимаются без необратимых изменений ультраструктуры протопласта. Сине‑зеленые водоросли (цианобактерии), вегетирующие на поверхности почвы в пустыне, высыхая, превращаются в темную корочку. От редких дождей их слизистая масса набухает и нитчатые тела начинают вегетировать. Мхи, растущие на сухих скалах, стволах деревьев или на поверхности почвы лугов и степей (роды Thuidium, Tortula и др.), также могут сильно высыхать, не теряя жизнеспособности.
Пойкилогидричны пыльцевые зерна и зародыши в семенах растений.
Гомойогидрические растения способны поддерживать относительное постоянство обводненности тканей. К ним относят большинство высших наземных растений. Для них характерна крупная центральная вакуоль в клетках. Благодаря этому клетка всегда имеет запас воды и не так сильно зависит от изменчивых внешних условий. Кроме того, побеги покрыты с поверхности эпидермой с малопроницаемой для воды кутикулой, транспирация регулируется устьичным аппаратом, а хорошо развитая корневая система во время вегетации может непрерывно поглощать влагу из почвы. Однако способности растений, не выдерживающих высыхания, регулировать свой водный обмен различны. Среди них выделяют разные по экологии группы.
3.3. Влажность3.3.2. Экологические группы растений по отношению к воде